

la productividad del Océano se refiere en gran medida a la producción de materia orgánica por, «plantas suspendidas en el océano, la mayoría de las cuales son unicelulares., El fitoplancton son «fotoautótrofos», que cosechan luz para convertir carbono inorgánico en orgánico, y suministran este carbono orgánico a diversos» heterótrofos», organismos que obtienen su energía únicamente de la respiración de la materia orgánica. Los heterótrofos del océano abierto incluyen bacterias, así como «zooplancton» unicelular y multicelular más complejos (animales flotantes), «nekton» (organismos nadadores, incluidos peces y mamíferos marinos) y los «Bentos» (la comunidad de organismos del fondo marino).,
los muchos ciclos anidados de carbono asociados con la productividad de los océanos se revelan en las siguientes definiciones (Bender et al. 1987) (Figura 1). La » producción primaria bruta «(GPP) se refiere a la tasa total de producción de carbono orgánico por autótrofos, mientras que la» respiración » se refiere a la oxidación que produce energía del carbono orgánico de vuelta al dióxido de carbono. La «producción primaria neta» (NPP) es GPP menos la propia tasa de respiración de los autótrofos; es, por lo tanto, la tasa a la que el metabolismo completo del fitoplancton produce biomasa., La «producción secundaria» (SP) se refiere típicamente a la tasa de crecimiento de la biomasa heterotrófica. Solo una pequeña fracción de la materia orgánica ingerida por organismos heterótrofos se utiliza para crecer, la mayoría respirando de nuevo a carbono inorgánico disuelto y nutrientes que pueden ser reutilizados por autótrofos. Por lo tanto, el SP en el océano es pequeño en comparación con el NPP. Las pesquerías dependen del SP; por lo tanto, dependen tanto de la NPP como de la eficiencia con la que la materia orgánica se transfiere a la red alimentaria (es decir, la relación SP/NPP)., La «producción neta del ecosistema» (NEP) es GPP menos la respiración de todos los organismos en el ecosistema. El valor de la NEP depende de los límites definidos para el ecosistema. Si se considera el océano superficial iluminado por el sol hasta el nivel de luz del 1% (la» zona eufótica») en el transcurso de un año entero, entonces la NEP es equivalente al carbono orgánico particulado que se hunde en el interior del océano oscuro más el carbono orgánico disuelto que circula fuera de la zona eufótica., En este caso, la NEP también se conoce a menudo como» producción de exportación «(o» Nueva producción»(Dugdale & Goering 1967), como se analiza a continuación). En contraste, la NEP para todo el océano, incluyendo sus sedimentos poco profundos, es aproximadamente equivalente al entierro lento de materia orgánica en los sedimentos menos la tasa de materia orgánica que entra desde los continentes.,
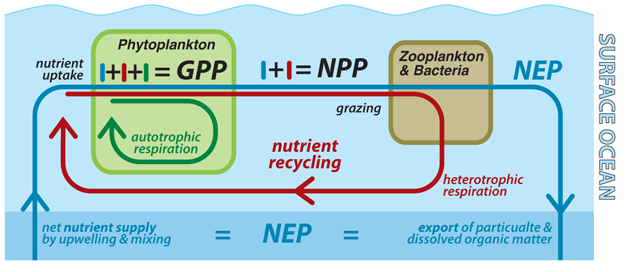
no Hay acumulaciones de biomasa viva en el medio marino que comparar con los bosques y pastizales en la tierra (Sarmiento & Bender 1994). Sin embargo, la biología oceánica es responsable del almacenamiento de más carbono lejos de la atmósfera que la biosfera terrestre (Broecker 1982)., Esto se logra mediante el hundimiento de la materia orgánica fuera de la superficie del océano y en el interior del océano antes de que sea devuelta al carbono inorgánico disuelto y a los nutrientes disueltos por descomposición bacteriana. Los oceanógrafos a menudo se refieren a este proceso como la «bomba biológica», ya que bombea dióxido de Carbono (CO2) fuera del océano superficial y la atmósfera y hacia el océano profundo voluminoso (Volk & Hoffert 1985).
solo una fracción de la materia orgánica producida en el océano superficial tiene el destino de ser exportada al océano profundo., De la materia orgánica producida por el fitoplancton (NPP), la mayor parte es respirada de nuevo a formas inorgánicas disueltas dentro de la superficie del océano y así reciclada para su uso por el fitoplancton (Eppley & Peterson 1979) (Figura 1). La mayoría de las células de fitoplancton son demasiado pequeñas para hundirse individualmente, por lo que el hundimiento ocurre solo una vez que se agregan en partículas más grandes o son empaquetadas en «gránulos fecales» por el zooplancton. Los restos de zooplancton también son suficientemente grandes para hundirse., Si bien el hundimiento es un destino relativamente raro para cualquier partícula dada en la superficie del océano, la biomasa y la materia orgánica no se acumulan en la superficie del océano, por lo que la exportación de materia orgánica por hundimiento es el destino final para todos los nutrientes que entran en la superficie del océano en forma disuelta, con las excepciones de que (1) los nutrientes disueltos pueden ser devueltos sin usar al interior por la circulación en algunas regiones polares (ver más abajo), y (2) la circulación también transporta la materia orgánica disuelta desde la superficie del océano al interior, un proceso significativo (Hansell et al., 2009) que no abordaremos más a fondo. A medida que la materia orgánica se asienta a través del interior del océano y en el fondo marino, se descompone casi por completo en productos químicos disueltos (Emerson & Hedges 2003, Martin et al. 1987). Esta alta eficiencia de descomposición se debe al hecho de que los organismos que llevan a cabo la descomposición dependen de ella como su única fuente de energía química; en la mayor parte del océano abierto, los heterótrofos solo dejan atrás la materia orgánica que es demasiado resistente químicamente para que valga la pena la inversión para descomponerse., En general, solo una pequeña fracción (típicamente mucho menos del 1%) del carbono orgánico de la central nuclear en la zona eufótica sobrevive para ser enterrado en sedimentos de aguas profundas.
la productividad en los ecosistemas costeros es a menudo distinta de la del océano abierto. A lo largo de las costas, el fondo marino es poco profundo, y la luz solar a veces puede penetrar a través de la columna de agua hasta el fondo, lo que permite que los organismos que habitan en el fondo («bentónicos») se fotosinteticen., Además, la materia orgánica que se hunde es percibida por el fondo marino, donde sustenta comunidades de fauna bentónica prósperas, en el proceso de ser reciclada de nuevo en nutrientes disueltos que luego están inmediatamente disponibles para la producción primaria. La proximidad a la tierra y sus fuentes de nutrientes, la interceptación de la materia orgánica que se hunde por el fondo marino poco profundo y la propensión a la surgencia costera dan como resultado ecosistemas altamente productivos. Aquí, nos referimos principalmente a la productividad del vasto océano abierto; sin embargo, muchos de los mismos conceptos, aunque en forma modificada, se aplican a los sistemas costeros.,
¿qué necesita la productividad Oceánica?
el fitoplancton requiere un conjunto de productos químicos, y aquellos con el potencial de ser escasos en las aguas superficiales se identifican típicamente como «nutrientes».»El calcio es un ejemplo de un elemento que es rápidamente asimilado por algunos plancton (para la producción de carbonato de calcio «partes duras»), pero no se considera típicamente un nutriente debido a su concentración uniformemente alta en el agua de mar. El carbono inorgánico disuelto, que es la materia prima para la producción de carbono orgánico por fotosíntesis, también es abundante y, por lo tanto, no suele figurar entre los nutrientes., Sin embargo, su forma ácida de CO2 disuelto es a menudo en concentraciones suficientemente bajas para afectar el crecimiento de al menos un poco de fitoplancton.
los nutrientes ampliamente importantes incluyen nitrógeno (N), fósforo (P), hierro (Fe) y silicio (Si). Parece que existen requisitos relativamente uniformes para el N y el P entre el fitoplancton. A principios de la década de 1900, el oceanógrafo Alfred Redfield descubrió que el plancton construye su biomasa con relaciones estequiométricas c:N:P de ~106:16:1, a las que ahora nos referimos como las relaciones de Redfield (Redfield 1958)., Como Redfield señaló, el n:P disuelto en el océano profundo está cerca de la relación de 16: 1 de la biomasa de plancton, y argumentaremos a continuación que el plancton impone esta relación en las profundidades, no viceversa. El hierro se encuentra en la biomasa solo en pequeñas cantidades, pero se utiliza para diversos propósitos esenciales en los organismos, y ha quedado claro en los últimos 25 años que la escasez de hierro a menudo limita o afecta la productividad en el océano abierto, especialmente en aquellas regiones donde las aguas profundas de alto N Y P son llevadas rápidamente a la superficie (Martin & Fitzwater 1988)., La investigación está en curso para comprender el papel de otros oligoelementos en la productividad (Morel et al. 2003). El silicio es un nutriente solo para taxones específicos del plancton-diatomeas (fitoplancton autotrófico), silicoflagelados y radiolaria (zooplancton heterotrófico) — que lo utilizan para hacer partes duras del ópalo. Sin embargo, el predominio típico de las diatomeas en las aguas que contienen Si, y la tendencia de la materia orgánica asociada a las diatomeas a hundirse fuera del océano superficial, hacen que la disponibilidad de Si sea un factor importante en la ecología y biogeoquímica más amplias de las aguas superficiales.,
La Luz Solar es la última fuente de energía — directa o indirectamente – para casi toda la vida en la tierra, incluso en el océano profundo. Sin embargo, la luz es absorbida y dispersada de tal manera que muy poca de ella penetra por debajo de una profundidad de ~80 m (tan profunda como 150 m en las regiones subtropicales menos productivas, pero tan poco profunda como 10 m en regiones altamente productivas y costeras) (Figura 2). Por lo tanto, la fotosíntesis se restringe en gran medida a la piel superior penetrada por la luz del Océano., Además, en la mayor parte del área del Océano, incluidos los trópicos, subtrópicos y la zona templada, la absorción de la luz solar hace que el agua superficial sea mucho más cálida que el océano profundo subyacente, este último lleno de agua que se hundió desde la superficie en las latitudes altas ., El agua caliente es más flotante que el frío, lo que hace que la capa superior iluminada por el sol flote en el océano profundo más denso, con la transición entre los dos conocida como la «picnoclina» (para «gradiente de densidad») o «termoclina» (el gradiente de temperatura vertical que impulsa la estratificación de densidad a través de la mayor parte del Océano, Figura 2). El viento u otra fuente de energía se requiere para conducir la mezcla a través de la picnoclina, por lo que el transporte de agua con sus productos químicos disueltos entre la superficie iluminada por el sol y el interior oscuro es lento., Este doble efecto de la luz sobre la fotosíntesis y la flotabilidad del agua de mar es fundamental para el éxito del fitoplancton oceánico. Si el océano no tuviera una fina capa superficial flotante, la mezcla sacaría a las algas de la luz y, por lo tanto, las alejaría de su fuente de energía la mayor parte del tiempo. En lugar de algas unicelulares casi neutralmente boyantes, organismos fotosintéticos más grandes y positivamente boyantes (por ejemplo, algas pelágicas) podrían dominar el océano abierto., Dejando de lado este caso hipotético, aunque se encuentran células de fitoplancton viables (aunque en bajas concentraciones) en aguas más profundas, la fotosíntesis limita el crecimiento activo del fitoplancton a la piel superior del océano, mientras que la estratificación de la densidad superior del Océano evita que se mezclen hacia el abismo oscuro. Por lo tanto, la mayor parte de la biomasa oceánica abierta, incluyendo fitoplancton, zooplancton y nekton, se encuentra dentro de ~200 m de la superficie del Océano.,

al mismo tiempo, la existencia de una capa superficial flotante delgada conspira con otros procesos para imponer limitaciones de nutrientes en la productividad del Océano. La exportación de materia orgánica a la profundidad agota el océano superficial de nutrientes, causando que los nutrientes se acumulen en aguas profundas donde no hay luz disponible para la fotosíntesis (Figura 2)., Debido a la diferencia de densidad entre el agua superficial y el mar profundo en la mayor parte del océano, la circulación oceánica solo puede reintroducir muy lentamente los nutrientes disueltos en la zona eufótica. Al expulsar los nutrientes de las aguas superficiales iluminadas por el sol y flotantes, la productividad del océano se limita efectivamente.
la limitación del crecimiento del fitoplancton se ha interpretado tradicionalmente en el contexto de la Ley del mínimo de Liebig, que establece que el crecimiento de las plantas será tan grande como lo permita el recurso menos disponible, el «nutriente limitante» que establece la productividad del sistema (de Baar 1994)., Si bien esta visión es poderosa, las interacciones entre los nutrientes y entre los nutrientes y la luz también pueden controlar la productividad. Un ejemplo simple pero importante de este potencial de «co-limitación» proviene de las regiones polares, donde la insolación solar oblicua se combina con una mezcla profunda de aguas superficiales para producir poca disponibilidad de luz. En tales ambientes, un mayor suministro de hierro puede aumentar la eficiencia con la que el fitoplancton captura la energía de la luz (Maldonado et al. 1999, Sunda & Huntsman 1997)., En términos más generales, se ha argumentado que el fitoplancton generalmente debe buscar un estado de co-limitación por todos los productos químicos que requieren, incluidos los muchos nutrientes metálicos traza (Morel 2008).
Deja una respuesta